第一节 细胞核的形态结构
Chapter 12 Interphase Nucleus and chromosomes
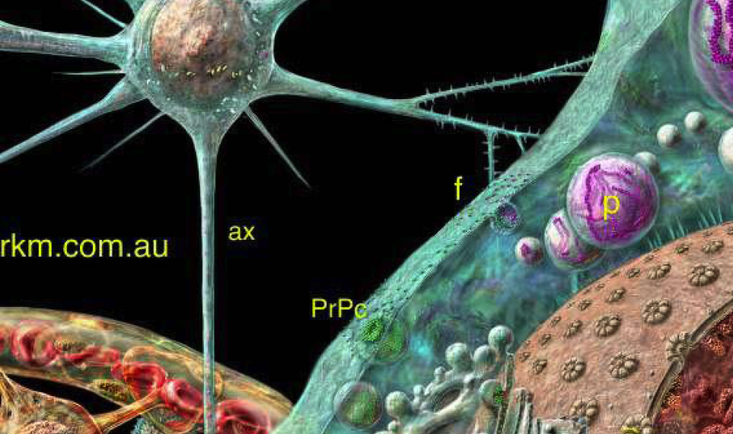
一般,真核细胞均具有细胞核,失去核后、很快就要死亡
特例: 哺乳动物的成熟红细胞(120 天) 植物韧皮部的筛管( 若干年)
细胞核数量:
一般,一个细胞只有一个细胞核
特殊细胞: 多核白细胞:多个核 ;纤毛虫:两个核( 生殖核,营养核)
细胞核的大小因物种不同而不同:
高等动物的细胞核:F 直径 10~15 mµ m
高等植物的细胞核:F 直径 5~20 mµ m
低等植物的细胞核:F 直径 1~4 mµ m
第一节 细胞核的形态结构
形状:球形或卵球形
物种和细胞类型不同
杆状 多叶状 枝状 带状
细胞核的形状往往会受到细胞形状的影响 ( 多角形、立方形和圆形细胞的核多呈圆形 )
细胞内含物的增多也会影响核的形状
细胞核多位于细胞的中央,但细胞内含物的增多可将核挤至细胞的一端。
间期核:核被膜 (保护核内遗传物质、控制核质之间的物质运输和信息交换) 、染色质 (遗传物质的存在形式) 、核仁 (核内浓密小体, 合成核糖体的场所) 、核质 (无定形结构的液体物质, 含有核体) 、核基质 (复杂的网络结构- 核骨架)、核孔(核质与胞质之间物质运输的通道)。
一、 核被膜与核孔复合体
一)核被膜 厚7.5nm
核孔 (nuclear pore) :内、外膜在一定部位相互融合而形成的环形开口
由100 多种蛋白质分子构成的分子量为0.5~1 ×105kDa 复杂结构——核孔复合体(pore complex)
孔膜区(pore membrane domain) :近核孔处的核膜,化学组成(其特征性蛋白为gp210 )
(二)核孔复合体——胞质与核质之间进行物质运输的通道
核孔复合体的数量随细胞种类不同而有较大差异:
在不同生理状态( 如基因转录活性) 下,核孔复合体的数量也会改变
1. 核孔复合体的结构
在脊椎动物中:约1.25 ´108 dalton( 核糖体的30 倍)
在酵母中:0.66 ´108 dalton
自1970 年Franke 、Roberts 等学者最早提出核孔复合体的模型以来,先后出现了多种模型。比较著名的有纤丝模型(Franke 和Scheer, 1974) 、滴漏样模型(H. Ris, 1991) 、圆柱模型(C.W. Akey, 1993) 和Ohno 模型(M. Ohno, 1998) 。
纤丝模型认为,核孔复合体是由核孔和孔环构成的直径80~120nm 的复合丝状结构,它在核膜上非随机分布,并穿越双层核膜形成通道。
首先,在核孔的内、外膜开口的边缘上均有环状物质存在,这些环状物质称为环带(annulus) 。环带为非匀质结构,核孔的内、外口处均有一个由8 个直径10~25nm 的环状颗粒(annular granules) 。核孔的中心还有一颗粒状或棒状的中央颗粒,称为中央栓(central plug) ,因它在核质交换中具有重要作用,故又被称为运输体(transporter) 。
每个环带的环状颗粒上还分别向核质与胞质中伸出丝状物。胞质环向胞质中伸出的丝状物短而卷曲;核质环向核质中伸出的丝状物长而直。
2. 核孔复合体的功能
(1) 核蛋白的运进
亲核蛋白(karyophilic protein) :核质蛋白(nucleoplasmin) :
一种酸性大的核蛋白,与核小体的装配有关
核输入信号(nuclear import signal)
又称为核定位信号(nuclear localization signal, NLS )
由4~8 个氨基酸构成的一段特殊的氨基酸序列,富含Lys 、Arg 和Pro 等带正电的氨基酸
此信号与信号肽不同:它可以定位在亲核蛋白的不同部位,而不仅仅是位于N- 末端;且在进入核后也不被切除.
亲核蛋白只有与受体蛋白结合后才能进入核内!
研究表明,亲核蛋白向细胞核输入的具体过程如下:
①亲核蛋白与输入蛋白α/ β异二聚体( 即NBP) 结合;
②形成的亲核蛋白- 受体复合体与核孔复合体的胞质丝结合;
③胞质丝向核弯曲,中央栓构象发生改变,形成亲水通道,蛋白质复合物进入核内;
④该复合体与Ran-GTP 相互作用,引起复合体解体,释放出亲核蛋白;
⑤输入蛋白β与Ran-GTP 结合在一起被运回细胞质,Ran-GTP 在细胞质中被水解为Ran-GDP ,Ran-GDP 随后被运回核内,而输入蛋白 α也在核内输出蛋白(exportin) 的帮助下运回细胞质。
(2) 核糖体蛋白的运进和核糖体亚单位(rRNP) 的运出
(3) 成熟mRNA 和scRNA 的运出
tRNA 的运出:也是一个蛋白质介导的过程
小 结
核孔复合体对亲核蛋白的运进和各种RNA 和核糖体亚单位的运出均具有高度选择性,运输过程既涉及主动运输又存在有被动运输。其主动运输是一个信号识别与载体介导的过程,需要消耗ATP 来提供能量。
(三)核纤层
位于内核膜内表面和染色质之间
由中间丝相互交织而形成的一层蛋白质网络结构
核纤层的厚度随细胞种类不同而有差异:
一般10~20nm ,最厚可30~100nm
核纤层的构件分子:——核纤层蛋白(lamins)
核纤层功能:
1. 维持核孔的位置和核被膜的形状;
2. 为间期染色质提供附着位点,是染色质的结构支架;
3. 在有丝分裂过程中,还与核被膜的解体和重建有关.
二、染色质和染色体的分子结构
一)染色质的分子结构
30nm 粗纤维为染色质的自然状态,10nm 细纤维为消化处理后产生的细线状结构
1. 染色质的基本结构单位-核小体(nucleosome)
由200 个碱基对的DNA 链结合有9 个组蛋白分子组成的重复亚单位.
(a) 核小体核心由146~147bp DNA 缠绕核心八聚体形成, DNA 的进头和出头为H1 所封闭构成了稳定的染色质小体, 含有165~166bp, 34~35bp 连接DNA 。(b) 核小体丝的模式图解。染色质小体靠连接区DNA 相互连接在一起,形成锯齿状的核小体丝结构。
DNA 复制时, 要发生核小体的局部解体和组蛋白8 聚体的全保留式组装
核酸酶的超敏感位点(nuclease hyper-sensitive site) :
易被DNase I 降解
非组蛋白等特异性DNA 结合蛋白的识别位点
基因表达的调控
2. 常染色质与异染色质
在间期和早前期中,染色质仍处于凝缩状态的那些区段
异染色质 :恒定型异染色质(constitutive heterochromatin) ,在各种细胞中总处于凝缩状态,最后进行复制。亦称组成型异染色质。
兼性异染色质(faculative heterochromatin) ,完全由异染色质组成的染色体( 如许多物种的性染色体) 。
二) 中期染色体的结构
1. 中期染色体结构的形成
非组蛋白在染色质结构中的组织作用
染色质折叠/ 压缩的分子机制:
间期染色质压缩形成中期染色体需要凝集蛋白(condensins) 的参与。
这类蛋白能利用ATP 水解释放的能量来驱动每条间期染色质的卷曲,进而形成一个分裂期染色体。
凝集蛋白是含有染色体结构维持(structural maintenance of chromosomes, SMC) 蛋白的大蛋白质复合体。SMC 蛋白是两个长蛋白分子在中心绞合形成的二聚体,其每个末端的球形结构域具有结合DNA 及水解ATP 的功能。如果将凝集蛋白加入到纯化DNA 中,凝集蛋白就能用ATP 水解能在DNA 上形成大的右手螺旋。
凝集蛋白在染色质压缩与折叠中的作用图解
(1) 着丝粒 (centromere)
(1) 着丝粒 (centromere)
位于染色体主缢痕处
着丝粒:
着丝粒DNA 序列
特异性着丝粒蛋白
动粒为动粒微管的组织中心:组织形成动粒微管,参与姊妹染色单体向细胞两极的分离
有些昆虫( 如蜜蜂、马蜂) 、某些显花植物( 如非洲紫罗兰) 和酵母没有明显的动粒结构,微管直接和染色质相连。
Plata(1990) 将哺乳动物细胞中染色体的主缢痕区域称为着丝粒- 动粒复合体,并提出了着丝粒- 动粒复合体的结构模型,将其分为动粒域(kinetochore domain) 、中心域(central domain) 和配对域(pairing domain) 三个结构域。
含有高度重复DNA ,对着丝粒—动粒复合体结构的形成和正常功能的维持有重要作用
动粒域与动粒微管连接,是支配染色体运动和分离的重要结构
为两条染色单体相互连接的区域,目前在此域中发现了内着丝粒蛋白INCENP 和染色单体连接蛋白CLIPs ,对染色体的配对连接和分离有重要作用
(2) 端粒(telomere)
由端粒DNA 与端粒蛋白构成
功能:
维持染色体的稳定性;
保证DNA 的完全复制;
参与染色体在核内的分布。
端粒酶的表达:端粒长:细胞分裂和存活 端粒短:不能分裂甚至不能存活
(3) 核仁组织区(nucleolus-organizing region, NOR)
位于次缢痕处
与核仁的形成有关 可活跃合成18S 、5.8S 和28SrRNA
核仁组织染色体:——拥有核仁组织区的染色体
人类有5 对同源染色体为核仁组织染色体
3. 染色体DNA 的关键序列 (key sequence)
自主复制DNA 序列(autonomously replicating sequence, ARS) : DNA 的自主复制;
着丝粒DNA 序列(centromere DNA sequence, CEN) :DNA 的平均分配;
端粒DNA 序列 (telomere DNA sequence, TEL) : DNA 复制的完整性。
4. 特殊染色体
(1) 多线染色体 (polytene chromosome)
多存在于双翅目昆虫( 如果蝇) 幼虫的唾腺细胞、肠细胞、气管细胞、马尔比基小管细胞以及植物的胚珠细胞中。
在双翅目昆虫的幼虫发育期间,染色体要进行4~15 次复制,复制后新、老链不分离,因而一个染色体中就有24~215 条DNA 分子,这种现象称为核内有丝分裂(endomitosis) 。
果蝇幼虫唾腺细胞核中的DNA 含量就比一般细胞多了1000 倍。这种只复制不分离的巨大染色体就称为多线染色体。由于多线染色体中平行排列的染色质纤维各段凝缩紧密程度不同,因而呈现出横带。
多线染色体的带可保持静息状态,但有的带也可发生蓬松现象,出现胀泡(puff) ( 旧名蓬突) ,是基因活动区,是正在进行转录的表现。转录的产物绝大部分是mRNA 。多线染色体的某些蓬突中也能掺入3H- 胸腺嘧啶核苷,说明这些区段进行DNA 外复制,是一种基因扩增现象。
最大的胀泡称为巴氏环。
(2) 灯刷染色体 (lampbrush chromosome)
动物卵母细胞在双线期中普遍发生染色体伸出大量侧环的现象。这时,染色体外形类似于灯刷( 或试管刷) ,故取名为灯刷染色体。
两栖类的灯刷染色体十分明显,而且很长,爪蟾的可长达5900 mm ,比果蝇的多线染色体长了3 倍。灯刷染色体出现于减数分裂的双线期,为二价体,两条同源染色体通过几处交叉而相连。每条染色体上可伸出5000~10000 个侧环。侧环是由DNA 分子外被以基质(matrix) 所组成,基质的成分是RNA 和蛋白质。每1个侧环是1个转录单位。
侧环两端粗细不等,由细端逐渐向另一端增粗。侧环上伸出的不同长度核糖核蛋白丝是DNA 轴的转录产物-mRNA 前体物,供卵细胞的生长和胚胎时期发育之用。
由于转录活动是由侧环一端开始,向着另一端进行,而且转录出的RNA 链与RNA 主轴垂直,因此随着DNA 链的不断延长而使侧环加粗,呈现为雪松形。
(3) B 染色体 (B chromosome)
1925 年,Kuwada 在玉米细胞中发现了一些二倍体之外的额外染色体,它们在形态、数目、行为及功能方面都与通常的染色体( 称为A 染色体) 不同。
Randolph(1928) 将这些染色体称为B 染色体。也称额外染色体(extra chr.) 、超数染色体(supernumerary chr.) 或附属染色体(accessory chr.) 。
B 染色体存在于自然界很多种动物和植物中。目前已在1000 多种植物及300 多种动物中发现了B 染色体。
B 染色体区别于A 染色体的显著特征
1. B 染色体通常比A 染色体小得多,且大多为端部或亚端部着丝粒,由大量的异染色质构成。
2. 它们与同一细胞中的任何一条A 染色体都不同源,且在减教分裂过程中只在B 染色体之间发生配对。
3. B 染色体的遗传不规则,一般不表现孟德尔式的遗传特征;当B 染色体数量较多时,可表现出数量遗传的特性。
4. 偶尔也会见到B 染色体可以形成核仁,但一般来说它们的遗传都是惰性的,其上不携带与主要性状相关的基因。
5. 在同一物种中,B 染色体也以多种形态出现。B 染色体的数目是可变的,在同一物种不同的组织、细胞、世代或个体之间,在有丝分裂和减数分裂中都常会有变化。
在植物对环境的适应上,B 染色体表现出积极的作用。在黑麦草属植物中,如果从播种到生长植株一直处于高密度,一部分植株就会被淘汰,而最后存活下来的往往是具有B 染色体的植株。
因此,在某些严峻的选择压力下,含有B 染色体的植株具有更强的竞争力和适应能力。
B 染色体对动植物的遗传、生长和发育具有多重效应,其起源、作用机制等问题都有待于进一步研究。
三、 核仁 (nucleolus)
真核细胞的间期核有1 个或几个浓密的球形小体,称为核仁(nucleolus) 。因为它较周围的核液(nucleoplasm) 浓密得多,故在光学显微镜下清晰可见。
核仁的形状、大小、数目,因生物种类和生理状态不同而有变动,一般生理活动旺盛的细胞,核仁大;不太活动的细胞,核仁就小。
核仁总是位于染色体组中一定染色体的特定部位,这种染色体称为核仁组织染色体(nucleolar organizing chromosome) 。
在电镜下可以看出,核仁无膜包围,它由四部分组成,即颗粒区(granular area) 、原纤维区(fibrilar area) 、核仁染色质和基质。其中,颗粒区和原纤维区最明显。
颗粒区往往位于核仁的边缘部分,是由直径约15~20nm 的颗粒( 略小于核糖体) 所组成。颗粒系处于不同成熟程度的核糖体亚单位前体。
原纤维区是由直径约为5~10nm 的原纤维所组成。用核糖核酸酶和蛋白质水解酶处理超薄切片,即显示出颗粒区和原纤维区显著减小,这说明此两种成分的性质为核糖核蛋白。
核仁染色质包括两部分,一部分围绕在核仁周围,称为核仁内染色质(intranucleolar chromotin) 。是含有核糖体DNA (rDNA) 的DNP 复合物。
核仁的前三种成分是悬浮在无结构的蛋白质性溶液中,此溶液即为核仁基质。
核仁四种成分的比例随细胞的生理活动而有所变化:细胞代谢旺盛时,颗粒区明显,反映出正在活跃地合成核糖体亚单位的状态。
核仁功能:
rRNA 转录与加工;
组装核糖体亚单位.
核仁功能活动的顺序:
纤维中心 致密纤维组分 颗粒组分
核仁在核糖体的合成与组装中的功能
核仁除了作为“核糖体工厂”之外,近年来的研究表明核仁还有其它一些重要的调节功能,是一种多功能的结构。
核仁调控许多细胞周期检验点控制蛋白的扣押(sequestration) ,如Mdm2( 与p53 连接) ,Cdc14 ( 一个促进细胞退出有丝分裂的蛋白磷酸酶) 和Pch2( 为利用监控点控制减数分裂周期进程暂停所必需) 。
此外的研究还表明,核仁在细胞增殖及存活方面也发挥着重要作用。
在热、电离辐射、拟辐射药物喜数碱等特定胁迫条件下,核仁又具有使核仁蛋白(nucleolin) 从核仁到核质重新定位的功能( 依赖于p53- 核仁蛋白复合体的形成) ,结果引起了核仁蛋白库的主体向核质的迁移,并在核质中与复制蛋白A(replication protein A, RPA) 相结合,从而抑制DNA 复制的启动。
此事件以及核仁ARF 蛋白( 有助于p53 稳定性) 对Mdm2 的扣押,表明了核仁在细胞反应胁迫中具有重要作用。
核仁的发生:分裂后期
核仁的发生:分裂后期,当rRNA 基因转录活动活跃时,围绕着由核仁组织区伸展开的rDNA 环形成了核仁。
每一物种均有一定的核仁组织染色体( 具有NOR) ,例如人类的为D 组的13 、14 、15 和G 组的21 和22 对染色体,小鼠的为12 、16 和18 对染色体。
一个物种的核仁数与核仁组织染色体的数目不一定一致,核仁往往相互融合在一起,如人的为1 个核仁,爪蟾为2 个核仁,摇蚊(Chironomus) 则为多个核仁。
四、核质及核体 (Nucleoplasm & Nuclear bodies)
1 、核质 (nucleoplasm): 间期细胞核中,在染色质和核仁周围的无定形的着色浅的液体。
经分析,这些颗粒和纤维状物质都是由RNA 复合以蛋白质所组成. 但其功能尚不完全清楚。
间期细胞核中,在染色质和核仁周围有无定形的着色浅的液体,称为核液(nucleoplasm) 。
在核液中,有一些颗粒状和细丝状物质。这些颗粒有两种,一种邻近染色质,直径40~45nm ,称为染色质周颗粒(perichromatin granules) ;另一种较小的颗粒,直径20~25 nm ,位于染色质之间。
此外还可以看到有染色质周纤维(perichromatin fiber) 和卷曲小体(coiled body) 。经化学分析,这些颗粒和纤维状物质都是RNP 。但它们的功能尚不清楚。
2 、核体 (Nuclear bodies, NBs):
核体是存在于核基质( 骨架) 中的一个异质性的核内( 核质) 结构集合体,主要是通过( 超微) 形态及抗原的标准来加以区分的。
核体包括含有球形和纤维状物质的多个结构域,排列成“卷曲小体”、囊、球或救生圈样的结构,偶尔也可见一些泡状或脂样结构。
一般认为核基质是核体的结构支架。
核体参与多种不同的生命活动:
如前体mRNA 的合成、加工、剪接、包装、核内转运、向核孔的运送以及向胞质的运输。
mRNA 及rRNA 的合成与加工过程,涉及到一些由不同RNA 及多种蛋白质组成的大型多元复合体,他们至少是部分出现在核体内的。
卷曲小体( coiled body, CBs) 最早由Ramóny Cajal 于1903 年在银染的神经元中发现。为常与核仁相连的Φ0.5 μm 的球形结构,因在TEM 下的结构类似于混乱的螺线卷,故而得名。
直到最近,才被正式命名为Cajal 小体,并发现其在结构上是由纤维状及球形结构组成的典型的薄片形卷曲结构( 直径0.1-0.2 μm) 。
Cajal 小体在多种不同的动植物细胞中均有发现,其数量与细胞周期进程、生理变化及疾病有关。
它们包含RNA 剪接所需要的许多因子。电镜免疫组化结果表明,Cajal 小体含有核质性的核小RNP (snRNPs) 、少量hnRNPs 、m3G- 帽snRNA 和核仁纤维蛋白(fibrillarin) 。
此外,一种80kDa 的卷曲蛋白(p80-coilin) 常被用作Cajal 小体的典型标记。
Cajal 小体是一种高度动态的结构,它们既能够相互聚集形成大的Cajal 小体,又能分解成可快速移动(0.9 μm/min) 的小Cajal 小体。
目前,Cajal 小体的功能还不甚清楚。但它却含有特异性核苷水解酶和snRNPs 、snRNAs ,故可能在转录后RNA 的代谢中具有积极作用。
因此,人们认为Cajal 小体极可能与snRNP 的贮存和/ 或前剪接复合体(pre-splicing complexes) 的装配有关。Cajal 小体很可能是作为一个含有snRNP 修饰酶的核内空间在起作用,可根据需要随时释放snRNP 至核仁中进行进一步修饰。
一些特定的癌基因编码蛋白可能也定位于癌细胞的Cajal 小体中,且在Cajal 小体中确实鉴定出了一种名为“pigpen ”的新的癌蛋白。
双卷曲小体(Gemini of coiled bodies, GEMs) 及染色质间颗粒簇( 又名小斑,speckle) 也是重要的核体。
双卷曲小体和Cajal 小体相互间很类似,且在核内常配对存在,至今还不清楚它们是否真的属于不同结构。Cajal 小体/ 双卷曲小体是snRNP 和snoRNP 的最终加工修饰部位。
相反,染色质间颗粒簇被认为是完全成熟snRNP 的贮存场所,以易于参加前体mRNA 的剪接。
第一节 细胞核的形态结构
Chapter 12 Interphase Nucleus and chromosomes
一般,真核细胞均具有细胞核,失去核后、很快就要死亡
特例: 哺乳动物的成熟红细胞(120 天) 植物韧皮部的筛管( 若干年)
细胞核数量:
一般,一个细胞只有一个细胞核
特殊细胞: 多核白细胞:多个核 ;纤毛虫:两个核( 生殖核,营养核)
细胞核的大小因物种不同而不同:
高等动物的细胞核:F 直径 10~15 mµ m
高等植物的细胞核:F 直径 5~20 mµ m
低等植物的细胞核:F 直径 1~4 mµ m
第一节 细胞核的形态结构
形状:球形或卵球形
物种和细胞类型不同
杆状 多叶状 枝状 带状
细胞核的形状往往会受到细胞形状的影响 ( 多角形、立方形和圆形细胞的核多呈圆形 )
细胞内含物的增多也会影响核的形状
细胞核多位于细胞的中央,但细胞内含物的增多可将核挤至细胞的一端。
间期核:核被膜 (保护核内遗传物质、控制核质之间的物质运输和信息交换) 、染色质 (遗传物质的存在形式) 、核仁 (核内浓密小体, 合成核糖体的场所) 、核质 (无定形结构的液体物质, 含有核体) 、核基质 (复杂的网络结构- 核骨架)、核孔(核质与胞质之间物质运输的通道)。
一、 核被膜与核孔复合体
一)核被膜 厚7.5nm
核孔 (nuclear pore) :内、外膜在一定部位相互融合而形成的环形开口
由100 多种蛋白质分子构成的分子量为0.5~1 ×105kDa 复杂结构——核孔复合体(pore complex)
孔膜区(pore membrane domain) :近核孔处的核膜,化学组成(其特征性蛋白为gp210 )
(二)核孔复合体——胞质与核质之间进行物质运输的通道
核孔复合体的数量随细胞种类不同而有较大差异:
在不同生理状态( 如基因转录活性) 下,核孔复合体的数量也会改变
1. 核孔复合体的结构
在脊椎动物中:约1.25 ´108 dalton( 核糖体的30 倍)
在酵母中:0.66 ´108 dalton
自1970 年Franke 、Roberts 等学者最早提出核孔复合体的模型以来,先后出现了多种模型。比较著名的有纤丝模型(Franke 和Scheer, 1974) 、滴漏样模型(H. Ris, 1991) 、圆柱模型(C.W. Akey, 1993) 和Ohno 模型(M. Ohno, 1998) 。
纤丝模型认为,核孔复合体是由核孔和孔环构成的直径80~120nm 的复合丝状结构,它在核膜上非随机分布,并穿越双层核膜形成通道。
首先,在核孔的内、外膜开口的边缘上均有环状物质存在,这些环状物质称为环带(annulus) 。环带为非匀质结构,核孔的内、外口处均有一个由8 个直径10~25nm 的环状颗粒(annular granules) 。核孔的中心还有一颗粒状或棒状的中央颗粒,称为中央栓(central plug) ,因它在核质交换中具有重要作用,故又被称为运输体(transporter) 。
每个环带的环状颗粒上还分别向核质与胞质中伸出丝状物。胞质环向胞质中伸出的丝状物短而卷曲;核质环向核质中伸出的丝状物长而直。
2. 核孔复合体的功能
(1) 核蛋白的运进
亲核蛋白(karyophilic protein) :核质蛋白(nucleoplasmin) :
一种酸性大的核蛋白,与核小体的装配有关
核输入信号(nuclear import signal)
又称为核定位信号(nuclear localization signal, NLS )
由4~8 个氨基酸构成的一段特殊的氨基酸序列,富含Lys 、Arg 和Pro 等带正电的氨基酸
此信号与信号肽不同:它可以定位在亲核蛋白的不同部位,而不仅仅是位于N- 末端;且在进入核后也不被切除.
亲核蛋白只有与受体蛋白结合后才能进入核内!
研究表明,亲核蛋白向细胞核输入的具体过程如下:
①亲核蛋白与输入蛋白α/ β异二聚体( 即NBP) 结合;
②形成的亲核蛋白- 受体复合体与核孔复合体的胞质丝结合;
③胞质丝向核弯曲,中央栓构象发生改变,形成亲水通道,蛋白质复合物进入核内;
④该复合体与Ran-GTP 相互作用,引起复合体解体,释放出亲核蛋白;
⑤输入蛋白β与Ran-GTP 结合在一起被运回细胞质,Ran-GTP 在细胞质中被水解为Ran-GDP ,Ran-GDP 随后被运回核内,而输入蛋白 α也在核内输出蛋白(exportin) 的帮助下运回细胞质。
(2) 核糖体蛋白的运进和核糖体亚单位(rRNP) 的运出
(3) 成熟mRNA 和scRNA 的运出
tRNA 的运出:也是一个蛋白质介导的过程
小 结
核孔复合体对亲核蛋白的运进和各种RNA 和核糖体亚单位的运出均具有高度选择性,运输过程既涉及主动运输又存在有被动运输。其主动运输是一个信号识别与载体介导的过程,需要消耗ATP 来提供能量。
(三)核纤层
位于内核膜内表面和染色质之间
由中间丝相互交织而形成的一层蛋白质网络结构
核纤层的厚度随细胞种类不同而有差异:
一般10~20nm ,最厚可30~100nm
核纤层的构件分子:——核纤层蛋白(lamins)
核纤层功能:
1. 维持核孔的位置和核被膜的形状;
2. 为间期染色质提供附着位点,是染色质的结构支架;
3. 在有丝分裂过程中,还与核被膜的解体和重建有关.
二、染色质和染色体的分子结构
一)染色质的分子结构
30nm 粗纤维为染色质的自然状态,10nm 细纤维为消化处理后产生的细线状结构
1. 染色质的基本结构单位-核小体(nucleosome)
由200 个碱基对的DNA 链结合有9 个组蛋白分子组成的重复亚单位.
(a) 核小体核心由146~147bp DNA 缠绕核心八聚体形成, DNA 的进头和出头为H1 所封闭构成了稳定的染色质小体, 含有165~166bp, 34~35bp 连接DNA 。(b) 核小体丝的模式图解。染色质小体靠连接区DNA 相互连接在一起,形成锯齿状的核小体丝结构。
DNA 复制时, 要发生核小体的局部解体和组蛋白8 聚体的全保留式组装
核酸酶的超敏感位点(nuclease hyper-sensitive site) :
易被DNase I 降解
非组蛋白等特异性DNA 结合蛋白的识别位点
基因表达的调控
2. 常染色质与异染色质
在间期和早前期中,染色质仍处于凝缩状态的那些区段
异染色质 :恒定型异染色质(constitutive heterochromatin) ,在各种细胞中总处于凝缩状态,最后进行复制。亦称组成型异染色质。
兼性异染色质(faculative heterochromatin) ,完全由异染色质组成的染色体( 如许多物种的性染色体) 。
二) 中期染色体的结构
1. 中期染色体结构的形成
非组蛋白在染色质结构中的组织作用
染色质折叠/ 压缩的分子机制:
间期染色质压缩形成中期染色体需要凝集蛋白(condensins) 的参与。
这类蛋白能利用ATP 水解释放的能量来驱动每条间期染色质的卷曲,进而形成一个分裂期染色体。
凝集蛋白是含有染色体结构维持(structural maintenance of chromosomes, SMC) 蛋白的大蛋白质复合体。SMC 蛋白是两个长蛋白分子在中心绞合形成的二聚体,其每个末端的球形结构域具有结合DNA 及水解ATP 的功能。如果将凝集蛋白加入到纯化DNA 中,凝集蛋白就能用ATP 水解能在DNA 上形成大的右手螺旋。
凝集蛋白在染色质压缩与折叠中的作用图解
(1) 着丝粒 (centromere)
(1) 着丝粒 (centromere)
位于染色体主缢痕处
着丝粒:
着丝粒DNA 序列
特异性着丝粒蛋白
动粒为动粒微管的组织中心:组织形成动粒微管,参与姊妹染色单体向细胞两极的分离
有些昆虫( 如蜜蜂、马蜂) 、某些显花植物( 如非洲紫罗兰) 和酵母没有明显的动粒结构,微管直接和染色质相连。
Plata(1990) 将哺乳动物细胞中染色体的主缢痕区域称为着丝粒- 动粒复合体,并提出了着丝粒- 动粒复合体的结构模型,将其分为动粒域(kinetochore domain) 、中心域(central domain) 和配对域(pairing domain) 三个结构域。
含有高度重复DNA ,对着丝粒—动粒复合体结构的形成和正常功能的维持有重要作用
动粒域与动粒微管连接,是支配染色体运动和分离的重要结构
为两条染色单体相互连接的区域,目前在此域中发现了内着丝粒蛋白INCENP 和染色单体连接蛋白CLIPs ,对染色体的配对连接和分离有重要作用
(2) 端粒(telomere)
由端粒DNA 与端粒蛋白构成
功能:
维持染色体的稳定性;
保证DNA 的完全复制;
参与染色体在核内的分布。
端粒酶的表达:端粒长:细胞分裂和存活 端粒短:不能分裂甚至不能存活
(3) 核仁组织区(nucleolus-organizing region, NOR)
位于次缢痕处
与核仁的形成有关 可活跃合成18S 、5.8S 和28SrRNA
核仁组织染色体:——拥有核仁组织区的染色体
人类有5 对同源染色体为核仁组织染色体
3. 染色体DNA 的关键序列 (key sequence)
自主复制DNA 序列(autonomously replicating sequence, ARS) : DNA 的自主复制;
着丝粒DNA 序列(centromere DNA sequence, CEN) :DNA 的平均分配;
端粒DNA 序列 (telomere DNA sequence, TEL) : DNA 复制的完整性。
4. 特殊染色体
(1) 多线染色体 (polytene chromosome)
多存在于双翅目昆虫( 如果蝇) 幼虫的唾腺细胞、肠细胞、气管细胞、马尔比基小管细胞以及植物的胚珠细胞中。
在双翅目昆虫的幼虫发育期间,染色体要进行4~15 次复制,复制后新、老链不分离,因而一个染色体中就有24~215 条DNA 分子,这种现象称为核内有丝分裂(endomitosis) 。
果蝇幼虫唾腺细胞核中的DNA 含量就比一般细胞多了1000 倍。这种只复制不分离的巨大染色体就称为多线染色体。由于多线染色体中平行排列的染色质纤维各段凝缩紧密程度不同,因而呈现出横带。
多线染色体的带可保持静息状态,但有的带也可发生蓬松现象,出现胀泡(puff) ( 旧名蓬突) ,是基因活动区,是正在进行转录的表现。转录的产物绝大部分是mRNA 。多线染色体的某些蓬突中也能掺入3H- 胸腺嘧啶核苷,说明这些区段进行DNA 外复制,是一种基因扩增现象。
最大的胀泡称为巴氏环。
(2) 灯刷染色体 (lampbrush chromosome)
动物卵母细胞在双线期中普遍发生染色体伸出大量侧环的现象。这时,染色体外形类似于灯刷( 或试管刷) ,故取名为灯刷染色体。
两栖类的灯刷染色体十分明显,而且很长,爪蟾的可长达5900 mm ,比果蝇的多线染色体长了3 倍。灯刷染色体出现于减数分裂的双线期,为二价体,两条同源染色体通过几处交叉而相连。每条染色体上可伸出5000~10000 个侧环。侧环是由DNA 分子外被以基质(matrix) 所组成,基质的成分是RNA 和蛋白质。每1个侧环是1个转录单位。
侧环两端粗细不等,由细端逐渐向另一端增粗。侧环上伸出的不同长度核糖核蛋白丝是DNA 轴的转录产物-mRNA 前体物,供卵细胞的生长和胚胎时期发育之用。
由于转录活动是由侧环一端开始,向着另一端进行,而且转录出的RNA 链与RNA 主轴垂直,因此随着DNA 链的不断延长而使侧环加粗,呈现为雪松形。
(3) B 染色体 (B chromosome)
1925 年,Kuwada 在玉米细胞中发现了一些二倍体之外的额外染色体,它们在形态、数目、行为及功能方面都与通常的染色体( 称为A 染色体) 不同。
Randolph(1928) 将这些染色体称为B 染色体。也称额外染色体(extra chr.) 、超数染色体(supernumerary chr.) 或附属染色体(accessory chr.) 。
B 染色体存在于自然界很多种动物和植物中。目前已在1000 多种植物及300 多种动物中发现了B 染色体。
B 染色体区别于A 染色体的显著特征
1. B 染色体通常比A 染色体小得多,且大多为端部或亚端部着丝粒,由大量的异染色质构成。
2. 它们与同一细胞中的任何一条A 染色体都不同源,且在减教分裂过程中只在B 染色体之间发生配对。
3. B 染色体的遗传不规则,一般不表现孟德尔式的遗传特征;当B 染色体数量较多时,可表现出数量遗传的特性。
4. 偶尔也会见到B 染色体可以形成核仁,但一般来说它们的遗传都是惰性的,其上不携带与主要性状相关的基因。
5. 在同一物种中,B 染色体也以多种形态出现。B 染色体的数目是可变的,在同一物种不同的组织、细胞、世代或个体之间,在有丝分裂和减数分裂中都常会有变化。
在植物对环境的适应上,B 染色体表现出积极的作用。在黑麦草属植物中,如果从播种到生长植株一直处于高密度,一部分植株就会被淘汰,而最后存活下来的往往是具有B 染色体的植株。
因此,在某些严峻的选择压力下,含有B 染色体的植株具有更强的竞争力和适应能力。
B 染色体对动植物的遗传、生长和发育具有多重效应,其起源、作用机制等问题都有待于进一步研究。
三、 核仁 (nucleolus)
真核细胞的间期核有1 个或几个浓密的球形小体,称为核仁(nucleolus) 。因为它较周围的核液(nucleoplasm) 浓密得多,故在光学显微镜下清晰可见。
核仁的形状、大小、数目,因生物种类和生理状态不同而有变动,一般生理活动旺盛的细胞,核仁大;不太活动的细胞,核仁就小。
核仁总是位于染色体组中一定染色体的特定部位,这种染色体称为核仁组织染色体(nucleolar organizing chromosome) 。
在电镜下可以看出,核仁无膜包围,它由四部分组成,即颗粒区(granular area) 、原纤维区(fibrilar area) 、核仁染色质和基质。其中,颗粒区和原纤维区最明显。
颗粒区往往位于核仁的边缘部分,是由直径约15~20nm 的颗粒( 略小于核糖体) 所组成。颗粒系处于不同成熟程度的核糖体亚单位前体。
原纤维区是由直径约为5~10nm 的原纤维所组成。用核糖核酸酶和蛋白质水解酶处理超薄切片,即显示出颗粒区和原纤维区显著减小,这说明此两种成分的性质为核糖核蛋白。
核仁染色质包括两部分,一部分围绕在核仁周围,称为核仁内染色质(intranucleolar chromotin) 。是含有核糖体DNA (rDNA) 的DNP 复合物。
核仁的前三种成分是悬浮在无结构的蛋白质性溶液中,此溶液即为核仁基质。
核仁四种成分的比例随细胞的生理活动而有所变化:细胞代谢旺盛时,颗粒区明显,反映出正在活跃地合成核糖体亚单位的状态。
核仁功能:
rRNA 转录与加工;
组装核糖体亚单位.
核仁功能活动的顺序:
纤维中心 致密纤维组分 颗粒组分
核仁在核糖体的合成与组装中的功能
核仁除了作为“核糖体工厂”之外,近年来的研究表明核仁还有其它一些重要的调节功能,是一种多功能的结构。
核仁调控许多细胞周期检验点控制蛋白的扣押(sequestration) ,如Mdm2( 与p53 连接) ,Cdc14 ( 一个促进细胞退出有丝分裂的蛋白磷酸酶) 和Pch2( 为利用监控点控制减数分裂周期进程暂停所必需) 。
此外的研究还表明,核仁在细胞增殖及存活方面也发挥着重要作用。
在热、电离辐射、拟辐射药物喜数碱等特定胁迫条件下,核仁又具有使核仁蛋白(nucleolin) 从核仁到核质重新定位的功能( 依赖于p53- 核仁蛋白复合体的形成) ,结果引起了核仁蛋白库的主体向核质的迁移,并在核质中与复制蛋白A(replication protein A, RPA) 相结合,从而抑制DNA 复制的启动。
此事件以及核仁ARF 蛋白( 有助于p53 稳定性) 对Mdm2 的扣押,表明了核仁在细胞反应胁迫中具有重要作用。
核仁的发生:分裂后期
核仁的发生:分裂后期,当rRNA 基因转录活动活跃时,围绕着由核仁组织区伸展开的rDNA 环形成了核仁。
每一物种均有一定的核仁组织染色体( 具有NOR) ,例如人类的为D 组的13 、14 、15 和G 组的21 和22 对染色体,小鼠的为12 、16 和18 对染色体。
一个物种的核仁数与核仁组织染色体的数目不一定一致,核仁往往相互融合在一起,如人的为1 个核仁,爪蟾为2 个核仁,摇蚊(Chironomus) 则为多个核仁。
四、核质及核体 (Nucleoplasm & Nuclear bodies)
1 、核质 (nucleoplasm): 间期细胞核中,在染色质和核仁周围的无定形的着色浅的液体。
经分析,这些颗粒和纤维状物质都是由RNA 复合以蛋白质所组成. 但其功能尚不完全清楚。
间期细胞核中,在染色质和核仁周围有无定形的着色浅的液体,称为核液(nucleoplasm) 。
在核液中,有一些颗粒状和细丝状物质。这些颗粒有两种,一种邻近染色质,直径40~45nm ,称为染色质周颗粒(perichromatin granules) ;另一种较小的颗粒,直径20~25 nm ,位于染色质之间。
此外还可以看到有染色质周纤维(perichromatin fiber) 和卷曲小体(coiled body) 。经化学分析,这些颗粒和纤维状物质都是RNP 。但它们的功能尚不清楚。
2 、核体 (Nuclear bodies, NBs):
核体是存在于核基质( 骨架) 中的一个异质性的核内( 核质) 结构集合体,主要是通过( 超微) 形态及抗原的标准来加以区分的。
核体包括含有球形和纤维状物质的多个结构域,排列成“卷曲小体”、囊、球或救生圈样的结构,偶尔也可见一些泡状或脂样结构。
一般认为核基质是核体的结构支架。
核体参与多种不同的生命活动:
如前体mRNA 的合成、加工、剪接、包装、核内转运、向核孔的运送以及向胞质的运输。
mRNA 及rRNA 的合成与加工过程,涉及到一些由不同RNA 及多种蛋白质组成的大型多元复合体,他们至少是部分出现在核体内的。
卷曲小体( coiled body, CBs) 最早由Ramóny Cajal 于1903 年在银染的神经元中发现。为常与核仁相连的Φ0.5 μm 的球形结构,因在TEM 下的结构类似于混乱的螺线卷,故而得名。
直到最近,才被正式命名为Cajal 小体,并发现其在结构上是由纤维状及球形结构组成的典型的薄片形卷曲结构( 直径0.1-0.2 μm) 。
Cajal 小体在多种不同的动植物细胞中均有发现,其数量与细胞周期进程、生理变化及疾病有关。
它们包含RNA 剪接所需要的许多因子。电镜免疫组化结果表明,Cajal 小体含有核质性的核小RNP (snRNPs) 、少量hnRNPs 、m3G- 帽snRNA 和核仁纤维蛋白(fibrillarin) 。
此外,一种80kDa 的卷曲蛋白(p80-coilin) 常被用作Cajal 小体的典型标记。
Cajal 小体是一种高度动态的结构,它们既能够相互聚集形成大的Cajal 小体,又能分解成可快速移动(0.9 μm/min) 的小Cajal 小体。
目前,Cajal 小体的功能还不甚清楚。但它却含有特异性核苷水解酶和snRNPs 、snRNAs ,故可能在转录后RNA 的代谢中具有积极作用。
因此,人们认为Cajal 小体极可能与snRNP 的贮存和/ 或前剪接复合体(pre-splicing complexes) 的装配有关。Cajal 小体很可能是作为一个含有snRNP 修饰酶的核内空间在起作用,可根据需要随时释放snRNP 至核仁中进行进一步修饰。
一些特定的癌基因编码蛋白可能也定位于癌细胞的Cajal 小体中,且在Cajal 小体中确实鉴定出了一种名为“pigpen ”的新的癌蛋白。
双卷曲小体(Gemini of coiled bodies, GEMs) 及染色质间颗粒簇( 又名小斑,speckle) 也是重要的核体。
双卷曲小体和Cajal 小体相互间很类似,且在核内常配对存在,至今还不清楚它们是否真的属于不同结构。Cajal 小体/ 双卷曲小体是snRNP 和snoRNP 的最终加工修饰部位。
相反,染色质间颗粒簇被认为是完全成熟snRNP 的贮存场所,以易于参加前体mRNA 的剪接。
第一节 细胞核的形态结构
Chapter 12 Interphase Nucleus and chromosomes
一般,真核细胞均具有细胞核,失去核后、很快就要死亡
特例: 哺乳动物的成熟红细胞(120 天) 植物韧皮部的筛管( 若干年)
细胞核数量:
一般,一个细胞只有一个细胞核
特殊细胞: 多核白细胞:多个核 ;纤毛虫:两个核( 生殖核,营养核)
细胞核的大小因物种不同而不同:
高等动物的细胞核:F 直径 10~15 mµ m
高等植物的细胞核:F 直径 5~20 mµ m
低等植物的细胞核:F 直径 1~4 mµ m
第一节 细胞核的形态结构
形状:球形或卵球形
物种和细胞类型不同
杆状 多叶状 枝状 带状
细胞核的形状往往会受到细胞形状的影响 ( 多角形、立方形和圆形细胞的核多呈圆形 )
细胞内含物的增多也会影响核的形状
细胞核多位于细胞的中央,但细胞内含物的增多可将核挤至细胞的一端。
间期核:核被膜 (保护核内遗传物质、控制核质之间的物质运输和信息交换) 、染色质 (遗传物质的存在形式) 、核仁 (核内浓密小体, 合成核糖体的场所) 、核质 (无定形结构的液体物质, 含有核体) 、核基质 (复杂的网络结构- 核骨架)、核孔(核质与胞质之间物质运输的通道)。
一、 核被膜与核孔复合体
一)核被膜 厚7.5nm
核孔 (nuclear pore) :内、外膜在一定部位相互融合而形成的环形开口
由100 多种蛋白质分子构成的分子量为0.5~1 ×105kDa 复杂结构——核孔复合体(pore complex)
孔膜区(pore membrane domain) :近核孔处的核膜,化学组成(其特征性蛋白为gp210 )
(二)核孔复合体——胞质与核质之间进行物质运输的通道
核孔复合体的数量随细胞种类不同而有较大差异:
在不同生理状态( 如基因转录活性) 下,核孔复合体的数量也会改变
1. 核孔复合体的结构
在脊椎动物中:约1.25 ´108 dalton( 核糖体的30 倍)
在酵母中:0.66 ´108 dalton
自1970 年Franke 、Roberts 等学者最早提出核孔复合体的模型以来,先后出现了多种模型。比较著名的有纤丝模型(Franke 和Scheer, 1974) 、滴漏样模型(H. Ris, 1991) 、圆柱模型(C.W. Akey, 1993) 和Ohno 模型(M. Ohno, 1998) 。
纤丝模型认为,核孔复合体是由核孔和孔环构成的直径80~120nm 的复合丝状结构,它在核膜上非随机分布,并穿越双层核膜形成通道。
首先,在核孔的内、外膜开口的边缘上均有环状物质存在,这些环状物质称为环带(annulus) 。环带为非匀质结构,核孔的内、外口处均有一个由8 个直径10~25nm 的环状颗粒(annular granules) 。核孔的中心还有一颗粒状或棒状的中央颗粒,称为中央栓(central plug) ,因它在核质交换中具有重要作用,故又被称为运输体(transporter) 。
每个环带的环状颗粒上还分别向核质与胞质中伸出丝状物。胞质环向胞质中伸出的丝状物短而卷曲;核质环向核质中伸出的丝状物长而直。
2. 核孔复合体的功能
(1) 核蛋白的运进
亲核蛋白(karyophilic protein) :核质蛋白(nucleoplasmin) :
一种酸性大的核蛋白,与核小体的装配有关
核输入信号(nuclear import signal)
又称为核定位信号(nuclear localization signal, NLS )
由4~8 个氨基酸构成的一段特殊的氨基酸序列,富含Lys 、Arg 和Pro 等带正电的氨基酸
此信号与信号肽不同:它可以定位在亲核蛋白的不同部位,而不仅仅是位于N- 末端;且在进入核后也不被切除.
亲核蛋白只有与受体蛋白结合后才能进入核内!
研究表明,亲核蛋白向细胞核输入的具体过程如下:
①亲核蛋白与输入蛋白α/ β异二聚体( 即NBP) 结合;
②形成的亲核蛋白- 受体复合体与核孔复合体的胞质丝结合;
③胞质丝向核弯曲,中央栓构象发生改变,形成亲水通道,蛋白质复合物进入核内;
④该复合体与Ran-GTP 相互作用,引起复合体解体,释放出亲核蛋白;
⑤输入蛋白β与Ran-GTP 结合在一起被运回细胞质,Ran-GTP 在细胞质中被水解为Ran-GDP ,Ran-GDP 随后被运回核内,而输入蛋白 α也在核内输出蛋白(exportin) 的帮助下运回细胞质。
(2) 核糖体蛋白的运进和核糖体亚单位(rRNP) 的运出
(3) 成熟mRNA 和scRNA 的运出
tRNA 的运出:也是一个蛋白质介导的过程
小 结
核孔复合体对亲核蛋白的运进和各种RNA 和核糖体亚单位的运出均具有高度选择性,运输过程既涉及主动运输又存在有被动运输。其主动运输是一个信号识别与载体介导的过程,需要消耗ATP 来提供能量。
(三)核纤层
位于内核膜内表面和染色质之间
由中间丝相互交织而形成的一层蛋白质网络结构
核纤层的厚度随细胞种类不同而有差异:
一般10~20nm ,最厚可30~100nm
核纤层的构件分子:——核纤层蛋白(lamins)
核纤层功能:
1. 维持核孔的位置和核被膜的形状;
2. 为间期染色质提供附着位点,是染色质的结构支架;
3. 在有丝分裂过程中,还与核被膜的解体和重建有关.
二、染色质和染色体的分子结构
一)染色质的分子结构
30nm 粗纤维为染色质的自然状态,10nm 细纤维为消化处理后产生的细线状结构
1. 染色质的基本结构单位-核小体(nucleosome)
由200 个碱基对的DNA 链结合有9 个组蛋白分子组成的重复亚单位.
(a) 核小体核心由146~147bp DNA 缠绕核心八聚体形成, DNA 的进头和出头为H1 所封闭构成了稳定的染色质小体, 含有165~166bp, 34~35bp 连接DNA 。(b) 核小体丝的模式图解。染色质小体靠连接区DNA 相互连接在一起,形成锯齿状的核小体丝结构。
DNA 复制时, 要发生核小体的局部解体和组蛋白8 聚体的全保留式组装
核酸酶的超敏感位点(nuclease hyper-sensitive site) :
易被DNase I 降解
非组蛋白等特异性DNA 结合蛋白的识别位点
基因表达的调控
2. 常染色质与异染色质
在间期和早前期中,染色质仍处于凝缩状态的那些区段
异染色质 :恒定型异染色质(constitutive heterochromatin) ,在各种细胞中总处于凝缩状态,最后进行复制。亦称组成型异染色质。
兼性异染色质(faculative heterochromatin) ,完全由异染色质组成的染色体( 如许多物种的性染色体) 。
二) 中期染色体的结构
1. 中期染色体结构的形成
非组蛋白在染色质结构中的组织作用
染色质折叠/ 压缩的分子机制:
间期染色质压缩形成中期染色体需要凝集蛋白(condensins) 的参与。
这类蛋白能利用ATP 水解释放的能量来驱动每条间期染色质的卷曲,进而形成一个分裂期染色体。
凝集蛋白是含有染色体结构维持(structural maintenance of chromosomes, SMC) 蛋白的大蛋白质复合体。SMC 蛋白是两个长蛋白分子在中心绞合形成的二聚体,其每个末端的球形结构域具有结合DNA 及水解ATP 的功能。如果将凝集蛋白加入到纯化DNA 中,凝集蛋白就能用ATP 水解能在DNA 上形成大的右手螺旋。
凝集蛋白在染色质压缩与折叠中的作用图解
(1) 着丝粒 (centromere)
(1) 着丝粒 (centromere)
位于染色体主缢痕处
着丝粒:
着丝粒DNA 序列
特异性着丝粒蛋白
动粒为动粒微管的组织中心:组织形成动粒微管,参与姊妹染色单体向细胞两极的分离
有些昆虫( 如蜜蜂、马蜂) 、某些显花植物( 如非洲紫罗兰) 和酵母没有明显的动粒结构,微管直接和染色质相连。
Plata(1990) 将哺乳动物细胞中染色体的主缢痕区域称为着丝粒- 动粒复合体,并提出了着丝粒- 动粒复合体的结构模型,将其分为动粒域(kinetochore domain) 、中心域(central domain) 和配对域(pairing domain) 三个结构域。
含有高度重复DNA ,对着丝粒—动粒复合体结构的形成和正常功能的维持有重要作用
动粒域与动粒微管连接,是支配染色体运动和分离的重要结构
为两条染色单体相互连接的区域,目前在此域中发现了内着丝粒蛋白INCENP 和染色单体连接蛋白CLIPs ,对染色体的配对连接和分离有重要作用
(2) 端粒(telomere)
由端粒DNA 与端粒蛋白构成
功能:
维持染色体的稳定性;
保证DNA 的完全复制;
参与染色体在核内的分布。
端粒酶的表达:端粒长:细胞分裂和存活 端粒短:不能分裂甚至不能存活
(3) 核仁组织区(nucleolus-organizing region, NOR)
位于次缢痕处
与核仁的形成有关 可活跃合成18S 、5.8S 和28SrRNA
核仁组织染色体:——拥有核仁组织区的染色体
人类有5 对同源染色体为核仁组织染色体
3. 染色体DNA 的关键序列 (key sequence)
自主复制DNA 序列(autonomously replicating sequence, ARS) : DNA 的自主复制;
着丝粒DNA 序列(centromere DNA sequence, CEN) :DNA 的平均分配;
端粒DNA 序列 (telomere DNA sequence, TEL) : DNA 复制的完整性。
4. 特殊染色体
(1) 多线染色体 (polytene chromosome)
多存在于双翅目昆虫( 如果蝇) 幼虫的唾腺细胞、肠细胞、气管细胞、马尔比基小管细胞以及植物的胚珠细胞中。
在双翅目昆虫的幼虫发育期间,染色体要进行4~15 次复制,复制后新、老链不分离,因而一个染色体中就有24~215 条DNA 分子,这种现象称为核内有丝分裂(endomitosis) 。
果蝇幼虫唾腺细胞核中的DNA 含量就比一般细胞多了1000 倍。这种只复制不分离的巨大染色体就称为多线染色体。由于多线染色体中平行排列的染色质纤维各段凝缩紧密程度不同,因而呈现出横带。
多线染色体的带可保持静息状态,但有的带也可发生蓬松现象,出现胀泡(puff) ( 旧名蓬突) ,是基因活动区,是正在进行转录的表现。转录的产物绝大部分是mRNA 。多线染色体的某些蓬突中也能掺入3H- 胸腺嘧啶核苷,说明这些区段进行DNA 外复制,是一种基因扩增现象。
最大的胀泡称为巴氏环。
(2) 灯刷染色体 (lampbrush chromosome)
动物卵母细胞在双线期中普遍发生染色体伸出大量侧环的现象。这时,染色体外形类似于灯刷( 或试管刷) ,故取名为灯刷染色体。
两栖类的灯刷染色体十分明显,而且很长,爪蟾的可长达5900 mm ,比果蝇的多线染色体长了3 倍。灯刷染色体出现于减数分裂的双线期,为二价体,两条同源染色体通过几处交叉而相连。每条染色体上可伸出5000~10000 个侧环。侧环是由DNA 分子外被以基质(matrix) 所组成,基质的成分是RNA 和蛋白质。每1个侧环是1个转录单位。
侧环两端粗细不等,由细端逐渐向另一端增粗。侧环上伸出的不同长度核糖核蛋白丝是DNA 轴的转录产物-mRNA 前体物,供卵细胞的生长和胚胎时期发育之用。
由于转录活动是由侧环一端开始,向着另一端进行,而且转录出的RNA 链与RNA 主轴垂直,因此随着DNA 链的不断延长而使侧环加粗,呈现为雪松形。
(3) B 染色体 (B chromosome)
1925 年,Kuwada 在玉米细胞中发现了一些二倍体之外的额外染色体,它们在形态、数目、行为及功能方面都与通常的染色体( 称为A 染色体) 不同。
Randolph(1928) 将这些染色体称为B 染色体。也称额外染色体(extra chr.) 、超数染色体(supernumerary chr.) 或附属染色体(accessory chr.) 。
B 染色体存在于自然界很多种动物和植物中。目前已在1000 多种植物及300 多种动物中发现了B 染色体。
B 染色体区别于A 染色体的显著特征
1. B 染色体通常比A 染色体小得多,且大多为端部或亚端部着丝粒,由大量的异染色质构成。
2. 它们与同一细胞中的任何一条A 染色体都不同源,且在减教分裂过程中只在B 染色体之间发生配对。
3. B 染色体的遗传不规则,一般不表现孟德尔式的遗传特征;当B 染色体数量较多时,可表现出数量遗传的特性。
4. 偶尔也会见到B 染色体可以形成核仁,但一般来说它们的遗传都是惰性的,其上不携带与主要性状相关的基因。
5. 在同一物种中,B 染色体也以多种形态出现。B 染色体的数目是可变的,在同一物种不同的组织、细胞、世代或个体之间,在有丝分裂和减数分裂中都常会有变化。
在植物对环境的适应上,B 染色体表现出积极的作用。在黑麦草属植物中,如果从播种到生长植株一直处于高密度,一部分植株就会被淘汰,而最后存活下来的往往是具有B 染色体的植株。
因此,在某些严峻的选择压力下,含有B 染色体的植株具有更强的竞争力和适应能力。
B 染色体对动植物的遗传、生长和发育具有多重效应,其起源、作用机制等问题都有待于进一步研究。
三、 核仁 (nucleolus)
真核细胞的间期核有1 个或几个浓密的球形小体,称为核仁(nucleolus) 。因为它较周围的核液(nucleoplasm) 浓密得多,故在光学显微镜下清晰可见。
核仁的形状、大小、数目,因生物种类和生理状态不同而有变动,一般生理活动旺盛的细胞,核仁大;不太活动的细胞,核仁就小。
核仁总是位于染色体组中一定染色体的特定部位,这种染色体称为核仁组织染色体(nucleolar organizing chromosome) 。
在电镜下可以看出,核仁无膜包围,它由四部分组成,即颗粒区(granular area) 、原纤维区(fibrilar area) 、核仁染色质和基质。其中,颗粒区和原纤维区最明显。
颗粒区往往位于核仁的边缘部分,是由直径约15~20nm 的颗粒( 略小于核糖体) 所组成。颗粒系处于不同成熟程度的核糖体亚单位前体。
原纤维区是由直径约为5~10nm 的原纤维所组成。用核糖核酸酶和蛋白质水解酶处理超薄切片,即显示出颗粒区和原纤维区显著减小,这说明此两种成分的性质为核糖核蛋白。
核仁染色质包括两部分,一部分围绕在核仁周围,称为核仁内染色质(intranucleolar chromotin) 。是含有核糖体DNA (rDNA) 的DNP 复合物。
核仁的前三种成分是悬浮在无结构的蛋白质性溶液中,此溶液即为核仁基质。
核仁四种成分的比例随细胞的生理活动而有所变化:细胞代谢旺盛时,颗粒区明显,反映出正在活跃地合成核糖体亚单位的状态。
核仁功能:
rRNA 转录与加工;
组装核糖体亚单位.
核仁功能活动的顺序:
纤维中心 致密纤维组分 颗粒组分
核仁在核糖体的合成与组装中的功能
核仁除了作为“核糖体工厂”之外,近年来的研究表明核仁还有其它一些重要的调节功能,是一种多功能的结构。
核仁调控许多细胞周期检验点控制蛋白的扣押(sequestration) ,如Mdm2( 与p53 连接) ,Cdc14 ( 一个促进细胞退出有丝分裂的蛋白磷酸酶) 和Pch2( 为利用监控点控制减数分裂周期进程暂停所必需) 。
此外的研究还表明,核仁在细胞增殖及存活方面也发挥着重要作用。
在热、电离辐射、拟辐射药物喜数碱等特定胁迫条件下,核仁又具有使核仁蛋白(nucleolin) 从核仁到核质重新定位的功能( 依赖于p53- 核仁蛋白复合体的形成) ,结果引起了核仁蛋白库的主体向核质的迁移,并在核质中与复制蛋白A(replication protein A, RPA) 相结合,从而抑制DNA 复制的启动。
此事件以及核仁ARF 蛋白( 有助于p53 稳定性) 对Mdm2 的扣押,表明了核仁在细胞反应胁迫中具有重要作用。
核仁的发生:分裂后期
核仁的发生:分裂后期,当rRNA 基因转录活动活跃时,围绕着由核仁组织区伸展开的rDNA 环形成了核仁。
每一物种均有一定的核仁组织染色体( 具有NOR) ,例如人类的为D 组的13 、14 、15 和G 组的21 和22 对染色体,小鼠的为12 、16 和18 对染色体。
一个物种的核仁数与核仁组织染色体的数目不一定一致,核仁往往相互融合在一起,如人的为1 个核仁,爪蟾为2 个核仁,摇蚊(Chironomus) 则为多个核仁。
四、核质及核体 (Nucleoplasm & Nuclear bodies)
1 、核质 (nucleoplasm): 间期细胞核中,在染色质和核仁周围的无定形的着色浅的液体。
经分析,这些颗粒和纤维状物质都是由RNA 复合以蛋白质所组成. 但其功能尚不完全清楚。
间期细胞核中,在染色质和核仁周围有无定形的着色浅的液体,称为核液(nucleoplasm) 。
在核液中,有一些颗粒状和细丝状物质。这些颗粒有两种,一种邻近染色质,直径40~45nm ,称为染色质周颗粒(perichromatin granules) ;另一种较小的颗粒,直径20~25 nm ,位于染色质之间。
此外还可以看到有染色质周纤维(perichromatin fiber) 和卷曲小体(coiled body) 。经化学分析,这些颗粒和纤维状物质都是RNP 。但它们的功能尚不清楚。
2 、核体 (Nuclear bodies, NBs):
核体是存在于核基质( 骨架) 中的一个异质性的核内( 核质) 结构集合体,主要是通过( 超微) 形态及抗原的标准来加以区分的。
核体包括含有球形和纤维状物质的多个结构域,排列成“卷曲小体”、囊、球或救生圈样的结构,偶尔也可见一些泡状或脂样结构。
一般认为核基质是核体的结构支架。
核体参与多种不同的生命活动:
如前体mRNA 的合成、加工、剪接、包装、核内转运、向核孔的运送以及向胞质的运输。
mRNA 及rRNA 的合成与加工过程,涉及到一些由不同RNA 及多种蛋白质组成的大型多元复合体,他们至少是部分出现在核体内的。
卷曲小体( coiled body, CBs) 最早由Ramóny Cajal 于1903 年在银染的神经元中发现。为常与核仁相连的Φ0.5 μm 的球形结构,因在TEM 下的结构类似于混乱的螺线卷,故而得名。
直到最近,才被正式命名为Cajal 小体,并发现其在结构上是由纤维状及球形结构组成的典型的薄片形卷曲结构( 直径0.1-0.2 μm) 。
Cajal 小体在多种不同的动植物细胞中均有发现,其数量与细胞周期进程、生理变化及疾病有关。
它们包含RNA 剪接所需要的许多因子。电镜免疫组化结果表明,Cajal 小体含有核质性的核小RNP (snRNPs) 、少量hnRNPs 、m3G- 帽snRNA 和核仁纤维蛋白(fibrillarin) 。
此外,一种80kDa 的卷曲蛋白(p80-coilin) 常被用作Cajal 小体的典型标记。
Cajal 小体是一种高度动态的结构,它们既能够相互聚集形成大的Cajal 小体,又能分解成可快速移动(0.9 μm/min) 的小Cajal 小体。
目前,Cajal 小体的功能还不甚清楚。但它却含有特异性核苷水解酶和snRNPs 、snRNAs ,故可能在转录后RNA 的代谢中具有积极作用。
因此,人们认为Cajal 小体极可能与snRNP 的贮存和/ 或前剪接复合体(pre-splicing complexes) 的装配有关。Cajal 小体很可能是作为一个含有snRNP 修饰酶的核内空间在起作用,可根据需要随时释放snRNP 至核仁中进行进一步修饰。
一些特定的癌基因编码蛋白可能也定位于癌细胞的Cajal 小体中,且在Cajal 小体中确实鉴定出了一种名为“pigpen ”的新的癌蛋白。
双卷曲小体(Gemini of coiled bodies, GEMs) 及染色质间颗粒簇( 又名小斑,speckle) 也是重要的核体。
双卷曲小体和Cajal 小体相互间很类似,且在核内常配对存在,至今还不清楚它们是否真的属于不同结构。Cajal 小体/ 双卷曲小体是snRNP 和snoRNP 的最终加工修饰部位。
相反,染色质间颗粒簇被认为是完全成熟snRNP 的贮存场所,以易于参加前体mRNA 的剪接。
第一节 细胞核的形态结构
Chapter 12 Interphase Nucleus and chromosomes
一般,真核细胞均具有细胞核,失去核后、很快就要死亡
特例: 哺乳动物的成熟红细胞(120 天) 植物韧皮部的筛管( 若干年)
细胞核数量:
一般,一个细胞只有一个细胞核
特殊细胞: 多核白细胞:多个核 ;纤毛虫:两个核( 生殖核,营养核)
细胞核的大小因物种不同而不同:
高等动物的细胞核:F 直径 10~15 mµ m
高等植物的细胞核:F 直径 5~20 mµ m
低等植物的细胞核:F 直径 1~4 mµ m
第一节 细胞核的形态结构
形状:球形或卵球形
物种和细胞类型不同
杆状 多叶状 枝状 带状
细胞核的形状往往会受到细胞形状的影响 ( 多角形、立方形和圆形细胞的核多呈圆形 )
细胞内含物的增多也会影响核的形状
细胞核多位于细胞的中央,但细胞内含物的增多可将核挤至细胞的一端。
间期核:核被膜 (保护核内遗传物质、控制核质之间的物质运输和信息交换) 、染色质 (遗传物质的存在形式) 、核仁 (核内浓密小体, 合成核糖体的场所) 、核质 (无定形结构的液体物质, 含有核体) 、核基质 (复杂的网络结构- 核骨架)、核孔(核质与胞质之间物质运输的通道)。
一、 核被膜与核孔复合体
一)核被膜 厚7.5nm
核孔 (nuclear pore) :内、外膜在一定部位相互融合而形成的环形开口
由100 多种蛋白质分子构成的分子量为0.5~1 ×105kDa 复杂结构——核孔复合体(pore complex)
孔膜区(pore membrane domain) :近核孔处的核膜,化学组成(其特征性蛋白为gp210 )
(二)核孔复合体——胞质与核质之间进行物质运输的通道
核孔复合体的数量随细胞种类不同而有较大差异:
在不同生理状态( 如基因转录活性) 下,核孔复合体的数量也会改变
1. 核孔复合体的结构
在脊椎动物中:约1.25 ´108 dalton( 核糖体的30 倍)
在酵母中:0.66 ´108 dalton
自1970 年Franke 、Roberts 等学者最早提出核孔复合体的模型以来,先后出现了多种模型。比较著名的有纤丝模型(Franke 和Scheer, 1974) 、滴漏样模型(H. Ris, 1991) 、圆柱模型(C.W. Akey, 1993) 和Ohno 模型(M. Ohno, 1998) 。
纤丝模型认为,核孔复合体是由核孔和孔环构成的直径80~120nm 的复合丝状结构,它在核膜上非随机分布,并穿越双层核膜形成通道。
首先,在核孔的内、外膜开口的边缘上均有环状物质存在,这些环状物质称为环带(annulus) 。环带为非匀质结构,核孔的内、外口处均有一个由8 个直径10~25nm 的环状颗粒(annular granules) 。核孔的中心还有一颗粒状或棒状的中央颗粒,称为中央栓(central plug) ,因它在核质交换中具有重要作用,故又被称为运输体(transporter) 。
每个环带的环状颗粒上还分别向核质与胞质中伸出丝状物。胞质环向胞质中伸出的丝状物短而卷曲;核质环向核质中伸出的丝状物长而直。
2. 核孔复合体的功能
(1) 核蛋白的运进
亲核蛋白(karyophilic protein) :核质蛋白(nucleoplasmin) :
一种酸性大的核蛋白,与核小体的装配有关
核输入信号(nuclear import signal)
又称为核定位信号(nuclear localization signal, NLS )
由4~8 个氨基酸构成的一段特殊的氨基酸序列,富含Lys 、Arg 和Pro 等带正电的氨基酸
此信号与信号肽不同:它可以定位在亲核蛋白的不同部位,而不仅仅是位于N- 末端;且在进入核后也不被切除.
亲核蛋白只有与受体蛋白结合后才能进入核内!
研究表明,亲核蛋白向细胞核输入的具体过程如下:
①亲核蛋白与输入蛋白α/ β异二聚体( 即NBP) 结合;
②形成的亲核蛋白- 受体复合体与核孔复合体的胞质丝结合;
③胞质丝向核弯曲,中央栓构象发生改变,形成亲水通道,蛋白质复合物进入核内;
④该复合体与Ran-GTP 相互作用,引起复合体解体,释放出亲核蛋白;
⑤输入蛋白β与Ran-GTP 结合在一起被运回细胞质,Ran-GTP 在细胞质中被水解为Ran-GDP ,Ran-GDP 随后被运回核内,而输入蛋白 α也在核内输出蛋白(exportin) 的帮助下运回细胞质。
(2) 核糖体蛋白的运进和核糖体亚单位(rRNP) 的运出
(3) 成熟mRNA 和scRNA 的运出
tRNA 的运出:也是一个蛋白质介导的过程
小 结
核孔复合体对亲核蛋白的运进和各种RNA 和核糖体亚单位的运出均具有高度选择性,运输过程既涉及主动运输又存在有被动运输。其主动运输是一个信号识别与载体介导的过程,需要消耗ATP 来提供能量。
(三)核纤层
位于内核膜内表面和染色质之间
由中间丝相互交织而形成的一层蛋白质网络结构
核纤层的厚度随细胞种类不同而有差异:
一般10~20nm ,最厚可30~100nm
核纤层的构件分子:——核纤层蛋白(lamins)
核纤层功能:
1. 维持核孔的位置和核被膜的形状;
2. 为间期染色质提供附着位点,是染色质的结构支架;
3. 在有丝分裂过程中,还与核被膜的解体和重建有关.
二、染色质和染色体的分子结构
一)染色质的分子结构
30nm 粗纤维为染色质的自然状态,10nm 细纤维为消化处理后产生的细线状结构
1. 染色质的基本结构单位-核小体(nucleosome)
由200 个碱基对的DNA 链结合有9 个组蛋白分子组成的重复亚单位.
(a) 核小体核心由146~147bp DNA 缠绕核心八聚体形成, DNA 的进头和出头为H1 所封闭构成了稳定的染色质小体, 含有165~166bp, 34~35bp 连接DNA 。(b) 核小体丝的模式图解。染色质小体靠连接区DNA 相互连接在一起,形成锯齿状的核小体丝结构。
DNA 复制时, 要发生核小体的局部解体和组蛋白8 聚体的全保留式组装
核酸酶的超敏感位点(nuclease hyper-sensitive site) :
易被DNase I 降解
非组蛋白等特异性DNA 结合蛋白的识别位点
基因表达的调控
2. 常染色质与异染色质
在间期和早前期中,染色质仍处于凝缩状态的那些区段
异染色质 :恒定型异染色质(constitutive heterochromatin) ,在各种细胞中总处于凝缩状态,最后进行复制。亦称组成型异染色质。
兼性异染色质(faculative heterochromatin) ,完全由异染色质组成的染色体( 如许多物种的性染色体) 。
二) 中期染色体的结构
1. 中期染色体结构的形成
非组蛋白在染色质结构中的组织作用
染色质折叠/ 压缩的分子机制:
间期染色质压缩形成中期染色体需要凝集蛋白(condensins) 的参与。
这类蛋白能利用ATP 水解释放的能量来驱动每条间期染色质的卷曲,进而形成一个分裂期染色体。
凝集蛋白是含有染色体结构维持(structural maintenance of chromosomes, SMC) 蛋白的大蛋白质复合体。SMC 蛋白是两个长蛋白分子在中心绞合形成的二聚体,其每个末端的球形结构域具有结合DNA 及水解ATP 的功能。如果将凝集蛋白加入到纯化DNA 中,凝集蛋白就能用ATP 水解能在DNA 上形成大的右手螺旋。
凝集蛋白在染色质压缩与折叠中的作用图解
(1) 着丝粒 (centromere)
(1) 着丝粒 (centromere)
位于染色体主缢痕处
着丝粒:
着丝粒DNA 序列
特异性着丝粒蛋白
动粒为动粒微管的组织中心:组织形成动粒微管,参与姊妹染色单体向细胞两极的分离
有些昆虫( 如蜜蜂、马蜂) 、某些显花植物( 如非洲紫罗兰) 和酵母没有明显的动粒结构,微管直接和染色质相连。
Plata(1990) 将哺乳动物细胞中染色体的主缢痕区域称为着丝粒- 动粒复合体,并提出了着丝粒- 动粒复合体的结构模型,将其分为动粒域(kinetochore domain) 、中心域(central domain) 和配对域(pairing domain) 三个结构域。
含有高度重复DNA ,对着丝粒—动粒复合体结构的形成和正常功能的维持有重要作用
动粒域与动粒微管连接,是支配染色体运动和分离的重要结构
为两条染色单体相互连接的区域,目前在此域中发现了内着丝粒蛋白INCENP 和染色单体连接蛋白CLIPs ,对染色体的配对连接和分离有重要作用
(2) 端粒(telomere)
由端粒DNA 与端粒蛋白构成
功能:
维持染色体的稳定性;
保证DNA 的完全复制;
参与染色体在核内的分布。
端粒酶的表达:端粒长:细胞分裂和存活 端粒短:不能分裂甚至不能存活
(3) 核仁组织区(nucleolus-organizing region, NOR)
位于次缢痕处
与核仁的形成有关 可活跃合成18S 、5.8S 和28SrRNA
核仁组织染色体:——拥有核仁组织区的染色体
人类有5 对同源染色体为核仁组织染色体
3. 染色体DNA 的关键序列 (key sequence)
自主复制DNA 序列(autonomously replicating sequence, ARS) : DNA 的自主复制;
着丝粒DNA 序列(centromere DNA sequence, CEN) :DNA 的平均分配;
端粒DNA 序列 (telomere DNA sequence, TEL) : DNA 复制的完整性。
4. 特殊染色体
(1) 多线染色体 (polytene chromosome)
多存在于双翅目昆虫( 如果蝇) 幼虫的唾腺细胞、肠细胞、气管细胞、马尔比基小管细胞以及植物的胚珠细胞中。
在双翅目昆虫的幼虫发育期间,染色体要进行4~15 次复制,复制后新、老链不分离,因而一个染色体中就有24~215 条DNA 分子,这种现象称为核内有丝分裂(endomitosis) 。
果蝇幼虫唾腺细胞核中的DNA 含量就比一般细胞多了1000 倍。这种只复制不分离的巨大染色体就称为多线染色体。由于多线染色体中平行排列的染色质纤维各段凝缩紧密程度不同,因而呈现出横带。
多线染色体的带可保持静息状态,但有的带也可发生蓬松现象,出现胀泡(puff) ( 旧名蓬突) ,是基因活动区,是正在进行转录的表现。转录的产物绝大部分是mRNA 。多线染色体的某些蓬突中也能掺入3H- 胸腺嘧啶核苷,说明这些区段进行DNA 外复制,是一种基因扩增现象。
最大的胀泡称为巴氏环。
(2) 灯刷染色体 (lampbrush chromosome)
动物卵母细胞在双线期中普遍发生染色体伸出大量侧环的现象。这时,染色体外形类似于灯刷( 或试管刷) ,故取名为灯刷染色体。
两栖类的灯刷染色体十分明显,而且很长,爪蟾的可长达5900 mm ,比果蝇的多线染色体长了3 倍。灯刷染色体出现于减数分裂的双线期,为二价体,两条同源染色体通过几处交叉而相连。每条染色体上可伸出5000~10000 个侧环。侧环是由DNA 分子外被以基质(matrix) 所组成,基质的成分是RNA 和蛋白质。每1个侧环是1个转录单位。
侧环两端粗细不等,由细端逐渐向另一端增粗。侧环上伸出的不同长度核糖核蛋白丝是DNA 轴的转录产物-mRNA 前体物,供卵细胞的生长和胚胎时期发育之用。
由于转录活动是由侧环一端开始,向着另一端进行,而且转录出的RNA 链与RNA 主轴垂直,因此随着DNA 链的不断延长而使侧环加粗,呈现为雪松形。
(3) B 染色体 (B chromosome)
1925 年,Kuwada 在玉米细胞中发现了一些二倍体之外的额外染色体,它们在形态、数目、行为及功能方面都与通常的染色体( 称为A 染色体) 不同。
Randolph(1928) 将这些染色体称为B 染色体。也称额外染色体(extra chr.) 、超数染色体(supernumerary chr.) 或附属染色体(accessory chr.) 。
B 染色体存在于自然界很多种动物和植物中。目前已在1000 多种植物及300 多种动物中发现了B 染色体。
B 染色体区别于A 染色体的显著特征
1. B 染色体通常比A 染色体小得多,且大多为端部或亚端部着丝粒,由大量的异染色质构成。
2. 它们与同一细胞中的任何一条A 染色体都不同源,且在减教分裂过程中只在B 染色体之间发生配对。
3. B 染色体的遗传不规则,一般不表现孟德尔式的遗传特征;当B 染色体数量较多时,可表现出数量遗传的特性。
4. 偶尔也会见到B 染色体可以形成核仁,但一般来说它们的遗传都是惰性的,其上不携带与主要性状相关的基因。
5. 在同一物种中,B 染色体也以多种形态出现。B 染色体的数目是可变的,在同一物种不同的组织、细胞、世代或个体之间,在有丝分裂和减数分裂中都常会有变化。
在植物对环境的适应上,B 染色体表现出积极的作用。在黑麦草属植物中,如果从播种到生长植株一直处于高密度,一部分植株就会被淘汰,而最后存活下来的往往是具有B 染色体的植株。
因此,在某些严峻的选择压力下,含有B 染色体的植株具有更强的竞争力和适应能力。
B 染色体对动植物的遗传、生长和发育具有多重效应,其起源、作用机制等问题都有待于进一步研究。
三、 核仁 (nucleolus)
真核细胞的间期核有1 个或几个浓密的球形小体,称为核仁(nucleolus) 。因为它较周围的核液(nucleoplasm) 浓密得多,故在光学显微镜下清晰可见。
核仁的形状、大小、数目,因生物种类和生理状态不同而有变动,一般生理活动旺盛的细胞,核仁大;不太活动的细胞,核仁就小。
核仁总是位于染色体组中一定染色体的特定部位,这种染色体称为核仁组织染色体(nucleolar organizing chromosome) 。
在电镜下可以看出,核仁无膜包围,它由四部分组成,即颗粒区(granular area) 、原纤维区(fibrilar area) 、核仁染色质和基质。其中,颗粒区和原纤维区最明显。
颗粒区往往位于核仁的边缘部分,是由直径约15~20nm 的颗粒( 略小于核糖体) 所组成。颗粒系处于不同成熟程度的核糖体亚单位前体。
原纤维区是由直径约为5~10nm 的原纤维所组成。用核糖核酸酶和蛋白质水解酶处理超薄切片,即显示出颗粒区和原纤维区显著减小,这说明此两种成分的性质为核糖核蛋白。
核仁染色质包括两部分,一部分围绕在核仁周围,称为核仁内染色质(intranucleolar chromotin) 。是含有核糖体DNA (rDNA) 的DNP 复合物。
核仁的前三种成分是悬浮在无结构的蛋白质性溶液中,此溶液即为核仁基质。
核仁四种成分的比例随细胞的生理活动而有所变化:细胞代谢旺盛时,颗粒区明显,反映出正在活跃地合成核糖体亚单位的状态。
核仁功能:
rRNA 转录与加工;
组装核糖体亚单位.
核仁功能活动的顺序:
纤维中心 致密纤维组分 颗粒组分
核仁在核糖体的合成与组装中的功能
核仁除了作为“核糖体工厂”之外,近年来的研究表明核仁还有其它一些重要的调节功能,是一种多功能的结构。
核仁调控许多细胞周期检验点控制蛋白的扣押(sequestration) ,如Mdm2( 与p53 连接) ,Cdc14 ( 一个促进细胞退出有丝分裂的蛋白磷酸酶) 和Pch2( 为利用监控点控制减数分裂周期进程暂停所必需) 。
此外的研究还表明,核仁在细胞增殖及存活方面也发挥着重要作用。
在热、电离辐射、拟辐射药物喜数碱等特定胁迫条件下,核仁又具有使核仁蛋白(nucleolin) 从核仁到核质重新定位的功能( 依赖于p53- 核仁蛋白复合体的形成) ,结果引起了核仁蛋白库的主体向核质的迁移,并在核质中与复制蛋白A(replication protein A, RPA) 相结合,从而抑制DNA 复制的启动。
此事件以及核仁ARF 蛋白( 有助于p53 稳定性) 对Mdm2 的扣押,表明了核仁在细胞反应胁迫中具有重要作用。
核仁的发生:分裂后期
核仁的发生:分裂后期,当rRNA 基因转录活动活跃时,围绕着由核仁组织区伸展开的rDNA 环形成了核仁。
每一物种均有一定的核仁组织染色体( 具有NOR) ,例如人类的为D 组的13 、14 、15 和G 组的21 和22 对染色体,小鼠的为12 、16 和18 对染色体。
一个物种的核仁数与核仁组织染色体的数目不一定一致,核仁往往相互融合在一起,如人的为1 个核仁,爪蟾为2 个核仁,摇蚊(Chironomus) 则为多个核仁。
四、核质及核体 (Nucleoplasm & Nuclear bodies)
1 、核质 (nucleoplasm): 间期细胞核中,在染色质和核仁周围的无定形的着色浅的液体。
经分析,这些颗粒和纤维状物质都是由RNA 复合以蛋白质所组成. 但其功能尚不完全清楚。
间期细胞核中,在染色质和核仁周围有无定形的着色浅的液体,称为核液(nucleoplasm) 。
在核液中,有一些颗粒状和细丝状物质。这些颗粒有两种,一种邻近染色质,直径40~45nm ,称为染色质周颗粒(perichromatin granules) ;另一种较小的颗粒,直径20~25 nm ,位于染色质之间。
此外还可以看到有染色质周纤维(perichromatin fiber) 和卷曲小体(coiled body) 。经化学分析,这些颗粒和纤维状物质都是RNP 。但它们的功能尚不清楚。
2 、核体 (Nuclear bodies, NBs):
核体是存在于核基质( 骨架) 中的一个异质性的核内( 核质) 结构集合体,主要是通过( 超微) 形态及抗原的标准来加以区分的。
核体包括含有球形和纤维状物质的多个结构域,排列成“卷曲小体”、囊、球或救生圈样的结构,偶尔也可见一些泡状或脂样结构。
一般认为核基质是核体的结构支架。
核体参与多种不同的生命活动:
如前体mRNA 的合成、加工、剪接、包装、核内转运、向核孔的运送以及向胞质的运输。
mRNA 及rRNA 的合成与加工过程,涉及到一些由不同RNA 及多种蛋白质组成的大型多元复合体,他们至少是部分出现在核体内的。
卷曲小体( coiled body, CBs) 最早由Ramóny Cajal 于1903 年在银染的神经元中发现。为常与核仁相连的Φ0.5 μm 的球形结构,因在TEM 下的结构类似于混乱的螺线卷,故而得名。
直到最近,才被正式命名为Cajal 小体,并发现其在结构上是由纤维状及球形结构组成的典型的薄片形卷曲结构( 直径0.1-0.2 μm) 。
Cajal 小体在多种不同的动植物细胞中均有发现,其数量与细胞周期进程、生理变化及疾病有关。
它们包含RNA 剪接所需要的许多因子。电镜免疫组化结果表明,Cajal 小体含有核质性的核小RNP (snRNPs) 、少量hnRNPs 、m3G- 帽snRNA 和核仁纤维蛋白(fibrillarin) 。
此外,一种80kDa 的卷曲蛋白(p80-coilin) 常被用作Cajal 小体的典型标记。
Cajal 小体是一种高度动态的结构,它们既能够相互聚集形成大的Cajal 小体,又能分解成可快速移动(0.9 μm/min) 的小Cajal 小体。
目前,Cajal 小体的功能还不甚清楚。但它却含有特异性核苷水解酶和snRNPs 、snRNAs ,故可能在转录后RNA 的代谢中具有积极作用。
因此,人们认为Cajal 小体极可能与snRNP 的贮存和/ 或前剪接复合体(pre-splicing complexes) 的装配有关。Cajal 小体很可能是作为一个含有snRNP 修饰酶的核内空间在起作用,可根据需要随时释放snRNP 至核仁中进行进一步修饰。
一些特定的癌基因编码蛋白可能也定位于癌细胞的Cajal 小体中,且在Cajal 小体中确实鉴定出了一种名为“pigpen ”的新的癌蛋白。
双卷曲小体(Gemini of coiled bodies, GEMs) 及染色质间颗粒簇( 又名小斑,speckle) 也是重要的核体。
双卷曲小体和Cajal 小体相互间很类似,且在核内常配对存在,至今还不清楚它们是否真的属于不同结构。Cajal 小体/ 双卷曲小体是snRNP 和snoRNP 的最终加工修饰部位。
相反,染色质间颗粒簇被认为是完全成熟snRNP 的贮存场所,以易于参加前体mRNA 的剪接。