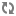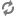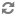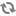第一节 质膜的分子结构
一、质膜的分子结构
质膜化学组成与分子结构方面的早期资料,主要来源于对哺乳动物血红细胞的研究。S.J. Singer和G.L. Nicolson在1972年提出了膜的流动镶嵌模型(fluid mosaic model):认为:构成膜的蛋白质和脂类分子具有镶嵌关系,而且膜的结构处于流动变化之中。
1、双层脂类分子构成了质膜的基本结构骨架
膜中的脂类分子为双性分子(有亲水头端和疏水尾端):亲水头端朝向水相,疏水尾端埋藏在膜的内部——呈双分子层排列,构成了膜的基本结构骨架。脂双分子层的内外两层是不对称的:在人RBC质膜中,卵磷脂(lecithin)(磷脂酰胆碱)和鞘磷脂(sphingomyelin)存在于脂双层的外单层。而含氨基的脂类,如磷脂酰乙醇胺(脑磷脂)、磷脂酰丝氨酸则主要存在于质膜脂双层的内单层。
2、膜的另一种主要成份是蛋白质
蛋白质分子或嵌插在脂双层网架中,或粘附在脂双层表面,根据在膜上的存在部位,膜蛋白可分为两类:一类蛋白质以不同深度嵌插在脂双层中,称为整合蛋白(integral protein)或内在蛋白(intrinsic protein)。另一类蛋白质附着于膜的表面,称为周边蛋白(peripheral protein)或外在蛋白(extrinsic protein)。
整合蛋白均为双性分子:其极性区(亲水区)朝向膜的表面,与水相接触;其非极性区(疏水区)则通过同脂双层脂分子的疏水尾部相互作用而插在脂双层之间(穿膜部分)。非极性区通过很强的疏水或亲水作用力同膜脂牢固结合,一般不易分离开来,除非采用破坏膜结构或使用去垢剂的强烈方法才能被提出。
周边蛋白与膜的结合比较疏松,用温和的方法即可将其分离下来。在人的红细胞内表面有一种红细胞膜蛋白——血影蛋白(spectrin)即为周边蛋白。
周边蛋白有的通过寡糖链与脂双层表面共价结合;有的则直接通过端部氨基酸残基与脂分子间共价结合;有的则附着在其它膜蛋白上。用冰冻蚀刻法可显示出蛋白质颗粒在脂双层中的分布状况。无论是整合蛋白还是周边蛋白,至少有一端露出膜表面,没有完全埋在膜内部的蛋白质分子。
流动镶嵌模型除了强调脂类分子与蛋白质分子的镶嵌关系外,还强调了膜的流动性。
主张:膜总是处于流动变化之中,脂类分子和蛋白质分子均可做侧向流动。
膜的流动性不是质膜所独有的属性,细胞内的各种膜,如内质网膜、线粒体膜等也都具有流动性。流动性也是生物膜的独有属性。
膜的特征可归纳为以下几点:
①镶嵌性:膜的基本结构是由脂双分子层镶嵌以蛋白质构成。双层脂分子以疏水尾相对,极性头朝向膜外水相;蛋白质则以不同程度镶嵌在脂双层中。
②流动性:构成膜的蛋白质分子和脂类分子在膜中的位置不断发生变化,脂类分子可发生侧向流动和倒翻等变化,蛋白质分子在膜中的位置亦可发生变动。
③不对称性:膜两侧的分子性质和结构不同,包括膜蛋白和膜脂在脂双层中的不对称分布。
④蛋白质极性:膜整合蛋白多肽链的极性区露出膜表面,而非极性区则埋在脂双层的内部,故蛋白质分子既与水溶性分子结合,也可与脂溶性分子亲和。
50年代末,伦敦大学的J.D. Robertson在电镜下发现,各种动物、植物和微生物细胞的质膜和各种细胞内膜在横切面上都呈“暗-明-暗”三层式结构。结合质膜的一些机能指标,他提出了单位膜模型。
指出单位膜为各种细胞的细胞膜以及各种细胞内膜在电镜下都呈“暗-明-暗”的三层式结构。
二、质膜的流动性
一)膜脂的流动性
膜脂流动性的大小对膜的功能活动,特别是酶的活性,具有重要影响。
1. 膜脂分子的运动方式:
①旋转异构运动 (烃链全反式构象-歪扭构象) ;
②左右摆动 (以与膜平面相垂直的轴线) ;
③旋转运动 (围绕与膜平面相垂直的轴) ;
④侧向扩散或侧向移动 (在膜内沿膜平面) ;
⑤倒翻运动 (两层脂分子中180°倒翻)。
2. 影响膜脂分子流动性的因素
①脂肪酸链的不饱和程度 (不饱和脂肪酸越多, 流动性越大)
②脂肪酸链的长度 (脂肪酸链短,相变温度低,流动性大)
③胆固醇/磷脂的比值 (胆固醇含量增加,膜脂流动性降低)
④卵磷脂/鞘磷脂比值 (鞘磷脂含量高,流动性低)
⑤膜蛋白的影响 (结合蛋白质,影响其流动性)
此外,膜脂的极性基团、环境温度、离子强度、金属离子等均可对膜脂的流动性产生一定的影响。
(二) 膜蛋白的运动性:
在各种情况下,膜蛋白均可发生位置上的变动。各种膜蛋白的运动速率不一样。
1、膜蛋白的运动方式:
侧向扩散:膜蛋白沿膜的二维表面运动;
旋转:绕与膜平面相垂直的轴线旋转。
2、膜蛋白的运动性的制约因素:
①周围膜脂的性质和相态:处于晶态脂质之滞流区中的膜蛋白不易运动;处于液态脂质区的膜蛋白则易于发生运动。
②质膜相关结构的作用:膜蛋白在膜中的运动并不是随脂质随机漂流,它还要受膜相关结构的影响。
③细胞骨架的作用:细胞质中的细胞骨架对膜蛋白的运动性具有动态控制作用(微管可固定膜蛋白的位置;而微丝可引起膜蛋白的运动)。在膜蛋白运动方面必然存在着一种跨膜的控制系统!
(三) 膜流动性的生理意义:
1. 膜蛋白酶活性:适宜的膜流动性是维持酶构象的必要条件;
2. 物质运输:载体蛋白, 受体介导内吞等均需要膜的流动性;
3. 细胞的信息传递:细胞的传递系统在膜上(激素及药物的作用需要适宜的膜流动性才能进行信号传递);
4. 细胞周期:分裂期最高, G1、S期最低;
5. 发育:一般, 成体细胞的膜流动性要小于幼体细胞; 经代谢来进行调节控制。(如贫血RBC低; 癌细胞高)
6. 植物耐寒性:与在低温下能否保持生物膜的流动性有关。
三、质膜的化学组成:
脂类——约占膜干重的30~70%
蛋白质——约占膜干重的20~70%
碳水化合物——约膜干重的10%
各种膜所含的蛋白质与脂类的比例大小同膜的功能有关(膜的功能主要由蛋白质承担):机能活动较旺盛的膜,蛋白质含量就高。
(一)膜脂
质膜的脂微区——脂筏:
脂分子在质膜上的分布不均匀,散布着一些脂微区(lipid microdomain),也称为脂筏(lipid raft)。脂筏主要是由鞘脂质(sphingolipid)、胆固醇和糖苷脂组成。由于鞘脂质的饱和脂尾比较长,因此由鞘脂质和胆固醇组成的脂筏比周围的脂双层要厚。
(二)膜蛋白
膜脂构成了膜的基本框架,但膜的许多功能是由膜蛋白来执行的——蛋白质是构成膜的另一种主要成分。
各种膜因性质和功能不同,其膜蛋白的含量有所不同(25%~80%)。
1、膜蛋白的存在形式
流动镶嵌模型最初将膜蛋白分为整合膜蛋白和周边膜蛋白两类。
现在根据蛋白质与质膜的结合关系,可将质膜蛋白质划分为三类:
①整合蛋白;②周边蛋白;③脂锚定蛋白。
三类膜蛋白:
①整合蛋白:均含有疏水区,该区与脂双层的疏水尾相结合。[疏水区一次或多次穿膜,穿膜片段往往是α螺旋,其它区段为β片层]
②周边蛋白:依靠电荷和氢键的相互作用,与整合蛋白或膜脂的极性头结合,从而附着在膜表面的蛋白质。 [此类蛋白由于不是共价键结合,故容易抽提出来 ]
③脂锚定蛋白:通过与脂锚形成共价键结合到膜上的蛋白质。蛋白质的一个氨基酸侧链通过酰胺键或脂键连接到豆蔻酸或棕榈酸的脂肪酰基上;脂肪酸插入到质膜的外脂层中。
整合蛋白12 周边蛋白3456
脂锚定蛋白:
脂锚定蛋白与质膜的结合方式
(a) 脂肪酰基锚定蛋白;(b) 异戊二烯锚定蛋白;(c) 糖基磷脂酰肌醇锚定蛋白
2. 整合蛋白的穿膜构象
跨膜蛋白,有的是一次穿膜,有的是多次穿膜,其穿膜部分的肽链大都以a-螺旋构象插在脂双层内部。
也有的跨膜蛋白是以β-折叠片构象穿膜,β-折叠片多次穿膜,并围成筒状结构,称为β筒(β barrel),例如孔蛋白(porin)的穿膜结构即是如此。